
Here’s the 2015 lab MUG designed by Silja Heilmann. Great for warm coffee or tea on a cloudy New York Sunday.

Here’s the 2015 lab MUG designed by Silja Heilmann. Great for warm coffee or tea on a cloudy New York Sunday.
Mathematical biology has always been obsessed with shapes, specially because many that look so complicated can actually be created with simple rules. Take the Turing patterns – these are regular shapes that obey the same type of rules but can make spots, stripes, labyrinths or holes by changing its parameters.

Turing-like patters in a model of biofilm, from Xavier et al (2009) Am Nat
Some years ago I read a paper by ecologists Rietkerk, Dekker, de Ruiter and van de Koppel that clicked for me. They explained that patterns in arid vegetation can have a simple explanation similar to Turing patterns. Plants in arid regions require moister (the growth limiting resource) and benefit from having other plants close by because neighbors provide shade and reduce water evaporation from the soil. However, too many neighbors means there is less moister to go around. In their words “vegetation patterns are the result of fine-scale positive feedback and coarse-scale negative feedback.” This means that having a few neighbors close by is good, but too many neighbors is bad. They presented a very simple cellular automata model that could reproduce the patterns. The cellular automaton is like a simple iterative game where the elements on a square grid follow a simple set of rules at every turn. Every element in the cellular automaton does this:
In the supporting material for their paper they show a convolution kernel:
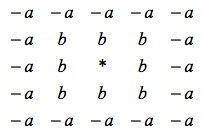
The kernel is an intuitive way to think about the cellular automaton rule. A focal bush located at the center of the kernel (represented by *) “looks” at the space around it and adds the value “b” or “-a” depending on whether that patch of land is occupied by another bush. If the final result is greater or equal to 0 then the focal bush gets to live and play another round. If the value is 0 then the bush dies and that grid element becomes free. The model is explained in detail the supporting material of the paper by Rietkerk et al. But the really striking result is that this very simple model produces a diversity of shapes depending on the value of a and b. The patterns can be made larger by using a larger kernel and also by changing the relative sizes of the “good” and “bad” neighborhoods.
We recently applied a similar idea to explain branching patterns of swarming colonies. Branching is a different process, we think, because it happens as a population spreads in space, but the patterns may have an equally simple explanation. In our case, a colony of Pseudomonas aeruginosa spreads across a petri dish and branches along the way suggesting that having too many neighbors is bad. We did some experiments that show support for these rules. For example, if we put two colonies in the same petri dish they repel each other suggesting the long range negative feedback (see video). Our model and experiments show that like the patterns in arid vegetation the branching in swarming can be caused by a short range positive feedback and a long range negative feedback (see video of model SIMSWARM).
Read the paper
The ecological basis of morphogenesis: branching patterns in swarming colonies of bacteria
Deng P, de Vargas Roditi L, van Ditmarsch D, Xavier JB. New Journal of Physics [Article: open access]
The paper has been selected to appear in the New Journal of Physics “Highlights of 2014” collection and recommended by Rob Palmer at the Faculty of 1000
Before the year ends I have to hurry and upload the pictures for our official 2014 X-Lab MUG.

Colonization resistance is the ability of the commensal microbiota to prevent invasion by entheric pathogens. This ability can be compromised when patients take antibiotics – a fact that was known for many antibiotics. However, the mechanisms by which colonization resistance works and why antibiotics compromise that resistance remains poorly understood.
In a paper that just came out today, we report n a significant step forward in understanding how the human gut microbiota protects against Clostridium difficile, an important human pathogen and problem specifically for hospitalized patients. The work was carried out in the Pamer lab. With Vanni Bucci and Richard Stein we analysed mouse and patient time series data to infer network models using the Lotka-Volterra method published last year.
Read our paper
Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile
Buffie CG et al. Nature
[Online]

Originally a meeting of Bacillus researchers located in the New York area, the NY/BIG has expanded to include other bacteria as well. This one day symposium also has speakers from beyond the NY area. Find more about it:
The primary function of intestinal microbiota seems to be to provide genes for metabolic reactions that are not available in the host’s genome. This enables us to process nutrients that would otherwise be unavailable to us. Another, perhaps secondary, function is to keep pathogens out. A healthy and biodiverse microbiota is resilient against invasion by invasive pathogenic bacteria such as Clostridium difficile and vancomycin-resistant Enterococcus (VRE). When we take antibiotics to cure an infection we may disrupt an ecological balance, compromising the microbiota’s resilience and opening the door for pathogens.
Microbiota resilience can involve ecological processes such as competition for nutrients, bacteriocin mediated bacterial warefare and the production of small molecules that stimulate the host to secrete antibacterial substances into the gut to harm competitor microbes. It should be possible to model these mechanisms with mathematics and build predictive computer models to help design antibiotic prescription regimens and minimize risk of disease.
In a review with Vanni Bucci we ask how far we are from such predictive models? Our conclusion is that we are probably far from a fully mechanistic, spatially structured model such as those used in environmental biotechnology. However, coarser grained models such as network inference and metabolic modeling are making great progress and may soon lead to the clinical applications.
This review is part of a special issue on the human microbiome in the Journal of Molecular Biology.

Microbiota dynamics of a bone marrow transplant patient
Read our review:
Towards predictive models of the human gut microbiome
Bucci V and Xavier JB. Journal of Molecular Biology [PDF]
How predictable can evolution be? This question is surely hard to answer. Even the smallest bacterial genomes have hundred of thousands of nucleotides that can be mutated and the genotypic space to explore can be huge. Moreover, even in a well controlled environment like a laboratory evolutionary experiment the selective pressures applied can be quite complex. Can we expect evolution to follow always the same path if we repeat experiments?
Recent work from our lab shows that the answer can be yes. We carried out experimental evolution using swarming motility in the bacterium Pseudomonas aeruginosa. Our experiments started with the wild-type strain PA14, which is mono-flagellated – it has a single polar flagellum. When we grow a colony of PA14 for 24 h on a Petri dish prepared with a minimal medium soft agar recipe, the colony forms a neat shape with a characteristic branched pattern. We passaged the bacteria to a new plate every day, and after only a few days we saw the bacteria evolve a different phenotype where they cover the entire plate. We call this phenotype “hyperswarming”.

The hyperswarmer evolution experiment
P. aeruginosa becomes a hyperswarmer by getting a point mutation in certain residues of a gene called fleN. The mutation makes the bacterium, which is normally mono-flagellated, become multi-flagellated.
Perhaps even more interesting, when we repeated the experiment several times we always found hyperswarmers with point mutations in the same gene. This is surprising because there are many genes and pathways known to affect swarming motility and therefore there were potentially many paths open for evolutionary adaptation.
Our experiments show that evolution can be, to some extent, predictable. The swarming experiments are based on a simple, well-established, microbiology protocol. In a recent article in Trends in Microbiology we encourage microbiologists to think about the evolutionary implications of their experiments.
Read also the piece by Carl Zimmer in his column “Matter” at the New York Times, which includes videos of swarming colonies.
Recent advances in DNA sequencing and metagenomics are opening a window into the human microbiome, and revealing novel associations between microbes, health and disease. But most microbiome studies are cross-sectional and lack a mechanistic understanding of this ecosystem.
We developed a method to analyze dynamics of microbiome composition which accounts for time-dependent external perturbations such as antibiotics. The new method combines Lotka–Volterra models of population dynamics with regression techniques and can be used to predict ecosystem dynamics.
We demonstrate the model using data from mouse experiments and we show that we can recover the microbiota temporal dynamics and study the concept of alternative stable states and antibiotic-induced transitions. The model suggests that a small group of commensal microbes protects against infection by the pathogen Clostridium difficile and explains how the antibiotic makes the host more susceptible to infection by perturbing the protective consortium.
Ecological modeling from time-series inference: insight into dynamics and stability of intestinal microbiota
Richard R. Stein*, Vanni Bucci*, Nora C. Toussaint, Charlie G. Buffie, Gunnar Rätsch, Eric G. Pamer, Chris Sander, João B. Xavier. PLoS Computational Biology
[Article: open access]
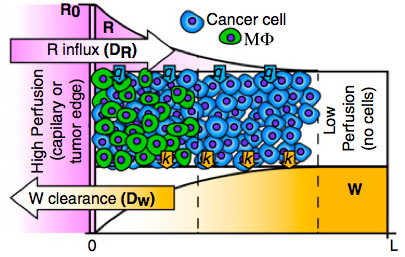
Lactic acid accumulation in the core of solid tumors can be harmful to macrophages
Cancer cells have dramatic metabolic alterations that can give growth advantages but also cause changes in the extracellular environment, i.e. the tumor microenvironment. We used a multidisciplinary combination of computational and experimental methods to show that lactic acid accumulation can impair the survival of tumor-associated macrophages. We show using in vitro models that the decreased survival can lead to spatial patterns of macrophage localization resembling how tumor-associated macrophages distribute in real tumors. Spatial patterns can potentiate tumor growth, and thus understanding how they are formed may bring therapeutic insights.
Emergence of spatial structure in the tumor microenvironment due to the Warburg effect
Carlos Carmona-Fontaine, Vanni Buccia, Leila Akkari, Maxime Deforet, J. A. Joyce, and Joao B. Xavier. PNAS [PDF]
Darwin’s theory of natural selection relies on the selfish survival of the fittest. Still, there are numerous examples of adaptive cooperative traits in nature. How can selection favor individuals that carry out costly actions for the benefit of others?
This question – how can cooperative traits evolve – is an old problem in evolutionary biology. Experimental microbiology is a growing source of models to tackle this problem.
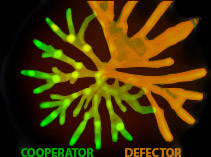
P. aeruginosa swarming: a model for the evolution of cooperation
In a recent article (Roditi et al, Molecular Systems Biology) we used swarming motility in the opportunistic pathogen Pseudomonas aeruginosa as a model for the evolution of cooperation. Laura Roditi, the first author, was the first PhD to graduate from our group and is now a postdoc with Manfred Claasen at the ETH in Zurich. The paper was also featured in a popular science article in the Brazilian science Instituto Ciencia Hoje (it’s in Portuguese, but a two year old can learn Portuguese).
Besides the fundamental implications to the evolution of cooperation, unveiling the mechanisms by which microbes stabilize cooperation can have implications to therapy – see this review:
Exploiting social evolution in biofilms
Boyle KE, Heilmann S, van Ditmarsch D, Xavier JB. Current Opinion in Microbiology.
